News & Announcements
Lipoamines
Article from 2018-01-22
aka Fatty Acid Amides; Lipo Amino Acids; N-acyl Amides; N-acyl Amino Acids
A novel lipid naming convention has recently been proposed by Dr. Heather Bradshaw, Associate Professor of Neuroscience at Indiana University Bloomington and the current president of The International Cannabinoid Research Society. Dr. Bradshaw has proposed the term lipoamine to encompass the family of lipids formed by the conjugation of fatty acids to any simple amine such as ethanolamine, monoamine neurotransmitters such as dopamine, taurine, serotonin, and GABA and all amino acids. These lipids are not novel per se. One of these compounds, N-arachidonoyl ethanolamine (anandamide) was identified as the first endocannabinoid; however, because of this there is an erroneous tendency to categorize all structurally analogous N-acyl ethanolamines as endocannabinoids. The new terminology LIPO + AMINE offers a simple and concise designation for any type of fatty acid amine conjugation through an amide bond (Figure 1).
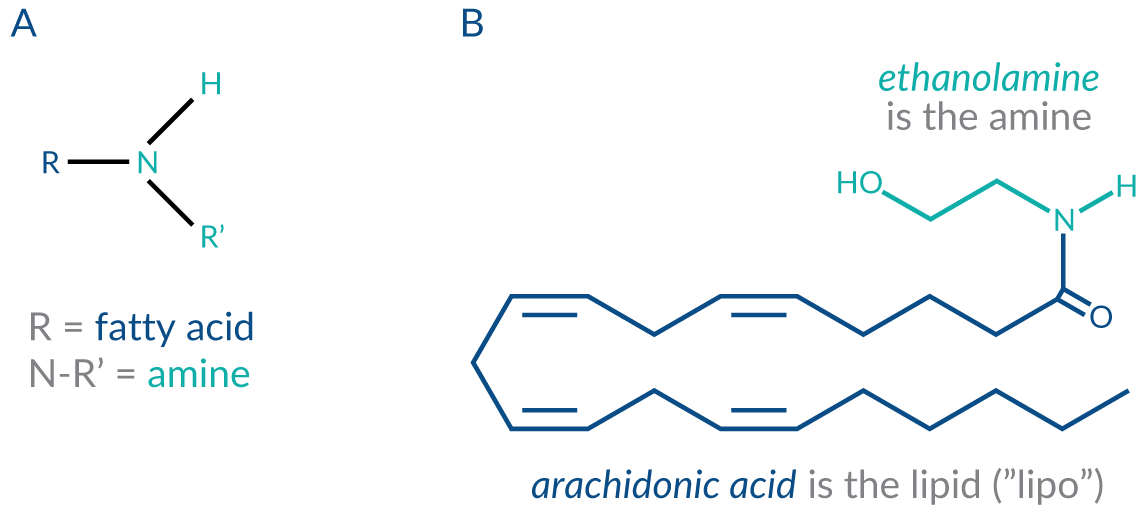
Figure 1. A. Structure of secondary amine. B. Anandamide is an archetype lipoamine.
This figure is adapted from Bradshaw, H.B. and Leishman, E. Lipidomics: A Corrective Lens for Enzyme Myopia. Methods Enzymol.593, 123-141 (2017).
Lipoamine-type conjugates include a subset of molecules that interact with a variety of other receptors, such as TRP channels and GPCRs, some of which are activated by cannabinoid compounds, but are not designated as cannabinoid receptors. Therefore, this terminology offers a means to avoid the assumption that if a fatty acid is conjugated to ethanolamine, then it must be an “endocannabinoid.”
Dr. Allyn Howlett, co-chair of the International Union Pharmacology Committee (IUPHAR) subcommittee on Cannabinoid Receptors, noted that “it is problematic to extend the name as an 'endocannabinoid' because that implies that whatever protein interacts with these lipoamines/lipoamides must also be a cannabinoid receptor. The IUPHAR Nomenclature Committee has determined that we do not wish to see that inference in research literature.”
We sat down to talk with Dr. Bradshaw about the new terminology.
What led you to propose the name lipoamines for this family of lipids?
The evolution of this nomenclature has been a long journey for me, and I will openly concede that it is likely far from over. It is ultimately driven by a desire to have a unified terminology that will allow all of those doing research on this class of compounds to be able to find each other’s work and grow within the field. These ideas have been building for years, but I will try to summarize how I got to this point. As a post-doc in Michael Walker’s lab starting in 2001, I had an opportunity to be at the forefront of the field of lipidomics and to begin isolating and characterizing novel lipids. When I started in Michael’s lab, we were only studying a few novel lipids, the first of which were N-arachidonoyl glycine or “NAGly” (we pronounced “nag-lie”) then N-arachidonoyl dopamine or “NADA,” and we simply called them by name. It wasn’t until we started to dig deeper and discover more structural analogs that the nomenclature began to be an issue. To give credit where it is due, it was Sumner Burstein from UMass Medical School in Worcester that had the original idea to look for these compounds in the brain and encouraged Michael to go in this direction because he had the tools when I joined his lab. Sumner was working on these compounds before either of us and was using the term “lipo amino acids,” which I liked and will come back around to. There was the understanding; however, that not all the amines of these conjugates were amino acids (e.g., ethanolamine). Sumner proposed the term “elmiric acids,” which as I understood was a combination of the first letters of the first names of each of his grandchildren. For many reasons, this nomenclature didn’t catch on, and we all started using the nomenclature of “N-acyl amides” in Michael’s lab and then in my own.
Throughout the years; however, I would get “corrected” by chemists especially who would let me know that this “N-acyl amide” nomenclature is incorrect in that structurally, these molecules are secondary amines. One of the final blows for the N-acyl amide nomenclature was from the “ChemNerds” at Wiki who completely trashed the named. One of my Honors students in 2015 wanted to make a Wiki page about the class of molecules and named the page “N-acyl amides.” When he submitted the page, it was instantly disputed by the “ChemNerds.” They stated that structurally, these are not amides and even went so far as to suggest that they were “N-acyl amines,” but admitted that they didn’t think that was “correct” nomenclature either. One of the two “ChemNerds” debating the entry stated that “I think this is some type of biochemistry nomenclature.” Clearly, there was no consensus. Around the same time, I was asked to chair a session on this emerging class of lipids for the British Pharmacological Society. The stipulation was that the discussion went beyond endocannabinoids, and I was asked to invite other experts in the field. I realized that I honestly didn’t know many (any) other people studying this class of lipids outside of the endocannabinoid researchers that I knew. When I looked up “N-acyl amides” the hits were primarily my work and almost no one else. I started to look up fatty acid amides and again, mostly work on the enzyme fatty acid amide hydrolase (FAAH) or those focused on anandamide. I thought about Sumner and looked up lipo amino acids and began to get a few more hits, and then I looked up some individual molecules by their structural names and found a few more people, which led me to other terms they used for this class of lipids. It became very clear that there is absolutely no consensus on a nomenclature, which is profoundly limiting our ability as a group to communicate with each other. In the end, the terms I found were “fatty acid amide,” “fatty acid amine,” “fatty acyl amide,” “N-acyl amide,” “N-acyl amine,” “N-acyl amino acids,” “lipid amides,” and “lipo amino acids,” though there are likely others. As stated before the “lipo amino acid” nomenclature was particularly appealing in its scope in that it is sufficiently broad to cover a huge percentage of these molecules, but it missed key members such as the ethanolamine, dopamine, and GABA conjugates. So, I reasoned that that since they are all amines structurally, why not simply call them “lipoamines” as an umbrella term to cover them all.
What techniques are you using to identify families of lipoamines and how do you characterize their function?
We use methanolic lipid extraction, C18 solid phase extraction partial purification to isolate the compounds and tandem mass spectrometry to identify lipoamines. Chloroform extractions are not ideal to isolate these lipids, so we use only methanol as our initial solvent. To date, we have identified them in every organism and tissue type that we have screened. Our primary limitation is a lack of standards for those that are theoretically present. We have generated conjugates from 6 different fatty acids and 17 different amines, however, since there are many more of these constituents in nature, there are very likely more of them to be discovered.
We have characterized function through receptor screening assays (GPCRs and TRPs to date) and lipidomics screening assays in tissue and plasma using pharmacological treatments, disease models, and knockout mice.
How many fatty acids and amine-containing compounds currently occupy the lists you have generated using your analytical techniques?
We maintain approximately 80 lipoamines in our library and have identified most of these in mammalian tissues. We have screened about 20 model organisms and always find a range of these, depending on the fatty acids that are present in the organism. There are ~5 that we do not typically detect and those are the conjugates of some of the least abundant fatty acids, so there is likely a detection limit we are reaching with our system.
How many potential lipoamines can theoretically be generated by combining the various fatty acids and amine compounds in these lists?
The short answer is that “it depends.” Essentially, I theorize that if an organism produces or uses (i.e., incorporates from the diet/environment) a fatty acid, that it has the probability/potential to conjugate that fatty acid with the amines that are present in that organism. So, these would be the limiting factors. In terms of numbers, this would mean that that depending on the organism it would be in the ~100 to ~400 different species.
Lipoamines are related to endocannabinoids via mutual biosynthetic and metabolic pathways. What differentiates lipoamines from endocannabinoids? What are their novel biological roles?
Importantly, there are only a few endogenous lipids that have been designated as endocannabinoids specifically anandamide (N-arachidonoyl ethanolamine) and 2-arachidonoyl glycerol. Arguably, there will be more as the field expands and as what is designated as a cannabinoid (e.g., cannabidiol, cannabigerol) also expands the number of endogenous counterparts will expand. So, the current relationship is that while one lipoamine, N-arachidonoyl ethanolamine, is an endocannabinoid, most other lipoamines are not endocannabinoids. As you point out, and our data support that, the enzymes that have been designated as those regulating the endocannabinoid system (FAAH, MAGL, NAPE-PLD) do play a role in regulating the wider lipoamine system; therefore, we need to reframe their activity as not just acting on the endocannabinoid system.
What are their novel biological roles? This is very much a work in progress. We have identified 20 of them that activate (either as agonists or antagonists) TRPV1-4 channels1 and have shown that NAGly and N-arachidonoyl serine have activity at GPR18.2-5 We have also shown that N-oleoyl serine is an endogenous signaling molecule in bone6,7 that can prevent symptoms of osteoporosis and is in high abundance in olive oil.8N-palmitoyl glycine drive calcium mobilization in neurons9 and disease models and knockouts1,6,7,10-22 show that these lipoamines are interconnected and likely have a broad range of signaling properties.
What is your opinion of how well the term lipoamines is being adopted and utilized in the research community at this point?
It’s gotten a very mixed reaction. In communities that this is a novel concept, it is accepted as a viable term and quickly embraced. In communities where there are individuals who have been using another term, most often one they too have generated, there is much more resistance. I understand that resistance. Having used N-acyl amides as my primary terminology for over a decade, I understand the desire to “stay the course” when building on your own work. My argument is not that “my” term should be adopted but that we need to adopt a nomenclature that both reflects the basic chemical structural nomenclature that has been established for decades and that has the potential to unify the field. Without a truly unifying nomenclature, we will all still be going in our own directions scientifically and “reinventing the wheel” because we missed foundational work from other groups. To miss out on another’s findings simply because we didn’t use the right search terms to find their work significantly impedes progress. As I stated at the beginning of the interview, I am open to ideas. I decided to make a stand that we need to deal with the nomenclature issue so that we can progress as a field and very much welcome all who want to join in the conversation.
All correspondence with Dr. Bradshaw can be directed to hbbradsh@indiana.edu.
References
1. Raboune, S., Stuart, J.M., Leishman, E., et al. Novel endogenous N-acyl amides activate TRPV1-4 receptors, BV-2 microglia, and are regulated in brain in an acute model of inflammation. Front. Cell Neurosci. 8, 195 (2014).
2. McHugh, D., Hu, S.S., Rimmerman, N., et al. N-arachidonoyl glycine, an abundant endogenous lipid, potently drives directed cellular migration through GPR18, the putative abnormal cannabidiol receptor. BMC Neurosci. 11, 44 (2010).
3. McHugh, D., Page, J., Dunn, E., et al. Δ9-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 165(8), 2414-2424 (2012).
4. McHugh, D., Roskowski, D., Xie, S., et al. Δ9-THC and N-arachidonoyl glycine regulate BV-2 microglial morphology and cytokine release plasticity: Implications for signaling at GPR18. Front. Pharmacol. 4, 162 (2013).
5. McHugh, D., Wager-Miller, J., Page, J., et al. siRNA knockdown of GPR18 receptors in BV-2 microglia attenuates N-arachidonoyl glycine-induced cell migration. J. Mol. Signal. 7(1), 10 (2012).
6. Bab, I., Smoum, R., Bradshaw, H., et al. Skeletal lipidomics: Regulation of bone metabolism by fatty acid amide family. Br. J. Pharmacol. 163(7), 1441-1446 (2011).
7. Smoum, R., Bar, A., Tan, B., et al. Oleoyl serine, an endogenous N-acyl amide, modulates bone remodeling and mass. Proc. Natl. Acad. Sci. U.S.A. 107(41), 17710-17715 (2010).
8. Bradshaw, H.B. and Leishman, E. Levels of bioactive lipids in cooking oils: Olive oil is the richest source of oleoyl serine. J. Basic Clin. Physiol. Pharmacol. 27(3), 247-252 (2016).
9. Rimmerman, N., Bradshaw, H.B., Hughes, H.V., et al. N-palmitoyl glycine, a novel endogenous lipid that acts as a modulator of calcium influx and nitric oxide production in sensory neurons. Mol. Pharmacol. 74(1), 213-224 (2008).
10. Koppel, J., Bradshaw, H., Goldberg, T.E., et al. Endocannabinoids in Alzheimer's disease and their impact on normative cognitive performance: A case-control and cohort study. Lipids Health Dis. 8, 2 (2009).
11. Hirota, Y., Daikoku, T., Tranguch, S., et al. Uterine-specific p53 deficiency confers premature uterine senescence and promotes preterm birth in mice. J. Clin. Invest. 120(3), 803-815 (2010).
12. Stuart, J.M., Paris, J.J., Frye, C., et al. Brain levels of prostaglandins, endocannabinoids, and related lipids are affected by mating strategies. Int. J. Endocrinol. 2013, 436252 (2013).
13. Tortoriello, G., Rhodes, B.P., Takacs, S.M., et al. Targeted lipidomics in Drosophila melanogaster identifies novel 2-monoacylglycerols and N-acyl amides. PLoS One 8(7), e67865 (2013).
14. Guindon, J., Lai, Y., Takacs, S.M., et al. Alterations in endocannabinoid tone following chemotherapy-induced peripheral neuropathy: Effects of endocannabinoid deactivation inhibitors targeting fatty-acid amide hydrolase and monoacylglycerol lipase in comparison to reference analgesics following cisplatin treatment. Pharmacol. Res. 67(1), 94-109 (2013).
15. Balakrishna, S., Song, W., Achanta, S., et al. TRPV4 inhibition counteracts edema and inflammation and improves pulmonary function and oxygen saturation in chemically induced acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 307(2), L158-L172 (2014).
16. Wu, C.S., Morgan, D., Jew, C.P., et al. Long-term consequences of perinatal fatty acid amino hydrolase inhibition. Br. J. Pharmacol. 171(6), 1420-1434 (2014).
17. Sałaga, M., Mokrowiecka, A., Zakrzewski, P.K., et al. Experimental colitis in mice is attenuated by changes in the levels of endocannabinoid metabolites induced by selective inhibition of fatty acid amide hydrolase (FAAH). J. Crohns Colitis 8(9), 998-1009 (2014).
18. Fichna, J., Sałaga, M., Stuart, J., et al. Selective inhibition of FAAH produces antidiarrheal and antinociceptive effect mediated by endocannabinoids and cannabinoid-like fatty acid amides. Neurogastroenterol. Motil. 26(4), 470-481 (2014).
19. Woolcott, O.O., Richey, J.M., Kabir, M., et al. High-fat diet-induced insulin resistance does not increase plasma anandamide levels or potentiate anandamide insulinotropic effect in isolated canine islets. PLoS One 10(4), e0123558 (2015).
20. Crowe, M.S., Leishman, E., Banks, M.L., et al. Combined inhibition of monoacylglycerol lipase and cyclooxygenases synergistically reduces neuropathic pain in mice. Br. J. Pharmacol. 172(7), 1700-1712 (2015).
21. Carey, L.M., Slivicki, R.A., Leishman, E., et al. A pro-nociceptive phenotype unmasked in mice lacking fatty-acid amide hydrolase. Mol. Pain 12 (2016).
22. Leishman, E., Mackie, K., Luquet, S., et al. Lipidomics profile of a NAPE-PLD KO mouse provides evidence of a broader role of this enzyme in lipid metabolism in the brain. Biochim. Biophys. Acta. 1861(6), 491-500 (2016).
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640