News & Announcements
Plasmalogens: Antioxidant, Signaling, and Structural Roles
Plasmalogens: Antioxidant, Signaling, and Structural Roles
Article from 2020-09-08
Plasmalogens are membrane glycerophospholipids that contain an acid labile vinyl ether group at the sn-1 position of the glycerol backbone, a fatty acid at the sn-2 position, and a polar head group at the sn-3 position (Figure 1). They were first named upon observation of a plasma aldehyde compound produced by acid treatment. As ether glycerophospholipids, they differ from the more common diacyl glycerophospholipids by having a fatty alcohol instead of a fatty acid at the sn-1 position. These fatty alcohols are typically saturated C16 (C16:0) or saturated and monounsaturated C18 (C18:0, C18:1) carbon chains, which are linked by a 1-0-(1Z-alkenyl) bond.
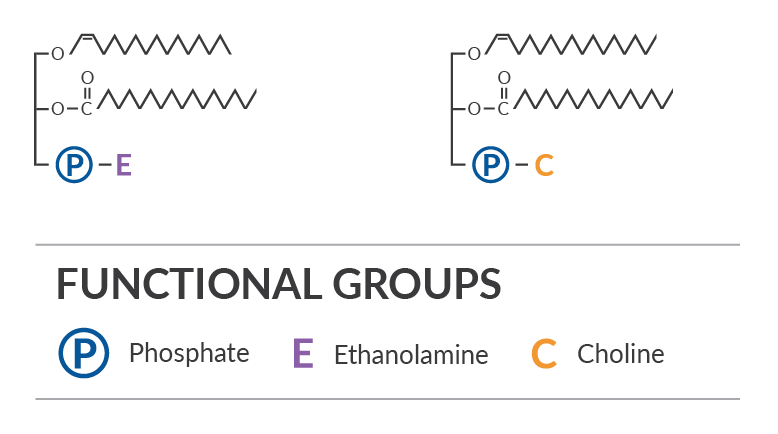
Plasmalogen Distribution, Synthesis, and Turnover
Plasmalogens comprise 15-20% of total phospholipids found in cell membranes. They are found in brain, heart, immune cells, kidney, lung, and skeletal muscle. They are synthesized in peroxisomes in the liver and transported by lipoproteins or through Golgi and endosomal vesicles to the membranes of tissues wherein their composition is controlled by remodeling, signaling induced hydrolysis, and degradation. Membrane plasmalogens can be remodeled by phospholipase A2 (PLA2) hydrolysis of the sn-2 group to remove the existing fatty acid and reacylation with a different fatty acid composition to produce a series of lipid messengers parallel to that of diacyl glycerophospholipids. Many of these fatty acids are introduced by exchange reactions from diacyl glycerophospholipids that are catalyzed by CoA-independent transacylases. Additionally, plasmalogens can be hydrolyzed by choline and ethanolamine glycerophospholipid-specific phospholipase C and phospholipase D. The ether bond in the sn-1 position of plasmalogens can be cleaved by cytochrome c, which is best known for its role in the mitochondria. Studies have shown that cytochrome c, which has been activated by cardiolipin to produce peroxidase activity, is released from myocardial mitochondria subjected to oxidative stress and will cleave plasmalogens in myocardium membranes. The end products of this peroxidase activity are a lysophospholipid and a highly reactive α-hydroxyaldehyde. The fatty alcohol on the sn-1 chain can also be oxidized by singlet oxygen, reactive oxygen species (ROS), or hypochlorous acid.
Plasmalogen Function
The sn-1 vinyl ether bond and the potential to be enriched with polyunsaturated fatty acids (PUFAs) at sn-2 position allows plasmalogens to have several functions unique from their diacyl glycerophospholipid counterparts. Structurally, their composition enables the sn-1 and sn-2 chains to remain in close apposition, which serves to decrease membrane fluidity, increase lateral domain organization, and facilitate lipid-protein interactions. Hydrophilicity is reduced by the lack of a carbonyl oxygen on the sn-1 chain. Plasmalogens are also highly concentrated in lipid rafts where they can influence membrane phase and signal transduction. Membranes of plasmalogen-deficient cells have higher lipid mobility, decreased order, and membrane receptor signaling defects. These deficient cells also display defects in cholesterol transport from the cell surface or endocytic compartments to the endoplasmic reticulum (ER) that reduce the pool of cholesterol available for efflux. As a result, plasmalogen-deficient cells have higher levels of membrane-free cholesterol.
Plasmalogens have been suggested to spare PUFAs and other vulnerable membrane lipids from oxidative damage. Indeed, singlet oxygen interacts more rapidly with the vinyl ether bond of ether lipids than with other lipids in vitro. There are also counter-suggestions that PUFAs, whose bis-allylic hydrogens allow easy hydrogen atom abstraction, protect plasmalogens against oxidative damage. The oxidative byproducts of plasmalogens are less toxic than the free aldehydes and hydroperoxides produced by oxidation at other unsaturated centers and are unable to further propagate lipid peroxidation. Cells that lack plasmalogens are more sensitive to chemical hypoxia, ROS, and cell death compared to wild-type cells.
By carrying important fatty acids at the sn-2 position, plasmalogens can serve as reservoirs for key lipid mediators and immunomodulators that could be released by PLA2 hydrolysis. This includes arachidonic acid, which is the precursor of prostaglandins, thromboxanes, and leukotrienes. Docosahexaenoic acid (DHA), which serve as the basis for the synthesis of the D-series resolvins, maresins, and neuroprotectins, is another key mediator associated with plasmalogens. Evidence in some tissues suggests that DHA is primarily targeted to plasmalogens, and this storage is enhanced through dietary DHA supplementation. Lysoplasmalogens are also produced from the PLA2 reaction and may have signaling functions. For example, lysoplasmenylcholine, acting as a second messenger, stimulates CREB phosphorylation leading to the activation of cAMP-dependent protein kinase. The plasmalogen form of phosphatidylethanolamine is a major precursor of the endocannabinoid anandamide in brain. Plasmenylcholine, found in heart mitochondria, may be important for the remodeling of cardiolipin.
Plasmalogen Deficiency and Disease
Defects in peroxisomal plasmalogen synthesis cause the malformation syndrome rhizomelic chondrodysplasia punctata (RCDP) and contribute to the pathology of Zellweger spectrum. Most RCDP cases are due to mutations in the gene encoding the peroxisomal protein transporter, PEX7, which is required for peroxisome localization of GNPAT and AGPS, the two peroxisomal enzymes required to initiate plasmalogen synthesis (Figure 2). The remainder are caused by defects in GNPAT and AGPS, the genes encoding these peroxisomal enzymes.
Figure 2. Plasmalogens are synthesized by the association of glyceronephosphate O-acyltransferase (GNPAT) and alkylglycerone phosphate synthase (AGPS) on the luminal side of the peroxisome membrane. The generation of the fatty alcohol by fatty alcohol reductase 1 (FAR1), which preferentially reduces C16 and C18 fatty acyl-CoAs, controls the rate of this reaction. Additional modifications needed to form mature plasmalogens occur in the ER and resemble those employed for diacyl glycerophospholipids.
The manifestation of RCDP demonstrates that plasmalogens are critical for brain, lens, cardiac, renal, pulmonary, and bone development and have sustained roles in life-long tissue maintenance. Respiratory disorders, neurodegenerative diseases, and inflammatory conditions have also been linked to plasmalogen deficiency. These conditions reveal the structural, antioxidant, and signaling roles of plasmalogens in normal physiology.
Plasmalogens are enriched on the edges of large and small airways, likely in pulmonary epithelial cells that require peroxisome metabolism and may serve a structural role in surfactant. Low plasmalogen levels in tracheal aspirates of premature infants correlate to increased risk of bronchopulmonary dysplasia. Surfactant preparations with high plasmalogen content has been shown to improve this respiratory issue. Asthma associated with ozone exposure may be caused by oxidation of surfactant lipids, as ozone selectively decreases plasmalogens levels relative to diacyl glycerophospholipids. Plasmalogen deficiency has also been linked to chronic obstructive pulmonary disease and is associated with smoking. Since the lung is a direct target of ROS, the antioxidant activity of plasmalogens might protect against respiratory disease.
The brain contains the highest distribution of plasmalogens, and reduction of these lipids is associated with Alzheimer’s disease, Parkinson's disease, Neimann-Pick type C, Down’s syndrome, and experimental autoimmune encephalomyelitis (a model of multiple sclerosis). As one example, plasmalogen deficiency, as well as generalized peroxisome dysfunction, may be a marker for Alzheimer’s disease pathology, though it is not known whether plasmalogen loss is a contributing cause or downstream effect. Increased oxidative damage and accumulation of α-hydroxyaldehydes, alterations in the availability of cholesterol and γ-secretase activity that can facilitate Aβ production, modulation of Aβ aggregation, depletion of neuroprotective lipids, alterations in cell proliferation signaling, and loss of gray matter support of synapse structure may all have plasmalogen-linked roles in disease progression. The presence of plasmalogens in some strains of gut microbiota is also being investigated in relation to the function of the microbiota-gut-brain axis in neurodegenerative diseases.
Decreased plasmalogen levels are also associated with chronic inflammation, metabolic syndrome, type II diabetes, and cardiovascular diseases. The increased lipid oxidation that accompanies these conditions may possibly deplete plasmalogen levels as they serve an antioxidant function. Their depletion would eventually leave other lipids vulnerable to oxidation, and this likely has structural consequences. Studies also suggest that changes in plasmalogen metabolism may contribute to the development of various types of cancer.
Plasmalogen Detection
Plasmalogen replacement therapy is actively being explored to correct some of these disease pathologies. However, these lipids present unique structural attributes in different tissue types, and their pathophysiological roles and possible metabolic pathways have not been easy to pinpoint. Further research must be done to determine the signature molecular species of plasmalogens associated with each different disease. By providing characteristic fragment ions for the head group class and fatty alcohols/acids esterified to the glycerol backbone, LC–MS/MS can facilitate the accurate identification of different plasmalogen subspecies, as well as novel plasmalogens that may have tissue specific functions. Cayman offers key plasmalogen standards to help gain broader knowledge about their activity both in normal physiology and disease.
Plasmalogen Standards
1-1(Z)-Hexadecenyl-2-Palmitoyl-sn-glycero-3-PE
1-1(Z)-Hexadecenyl-2-Palmitoyl-d9-sn-glycero-3-PE
1-1(Z)-Hexadecenyl-2-Palmitoyl-sn-glycero-3-PC
1-1(Z)-Hexadecenyl-2-Palmitoyl-d9-sn-glycero-3-PC
See all glycerophospholipids from Cayman
Lipidomics Services
Our lipid experts have decades of experience in lipid synthesis, purification, and characterization with access to state-of-the-art equipment for discovery and targeted lipidomics. Using an untargeted approach, our team can create a detailed lipid profile from thousands of lipid species or identify profile changes within a single lipid class. If you already know your lipid species of interest, our team can perform targeted lipidomics on a known collection of lipids utilizing our standardized protocols and established LC-MS/MS methods. Our approach can quantify lipids present at low levels in samples and reveal changing trends in lipid species amongst experimental groups using sophisticated data analysis software.
Custom Lipid Synthesis Services
Our growing team of more than 70 skilled and experienced chemists has broad synthetic proficiencies that include lipids, their bioconjugates, and stable isotope derivatives. We specialize in multi-step organic synthesis, purification, and characterization of complex fatty acid metabolites, glycerophospholipids, sphingolipids, and sterols.
Ready-to-Use, Quantitative MaxSpec® Standards
Cayman has translated its vast expertise in the synthesis, purification, and characterization of lipids into our line of MaxSpec® standards designed to simplify mass spectrometry workflows for prostaglandins, docosanoids, fatty acids, and more. MaxSpec® standards are gravimetrically prepared in solution and supplied in glass ampules to save time in the preparation of standards and samples. In addition to providing ease of use, MaxSpec® standards are characterized with a defined concentration as well as by purity and identity tests to enable quantitative accuracy and reproducibility.
You May Also Be Interested In
Suggested Reading
Braverman, N.E. and Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 1822(9), 1442-1452 (2012).
Dean, J.M. and Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell9(2), 196-206 (2018).
Dorninger, F., Forss-Petter, S., and Berger, J. From peroxisomal disorders to common neurodegenerative diseases - the role of ether phospholipids in the nervous system. FEBS Lett.591(18), 2761-2788 (2017).
Fontaine, D., Figiel, S., Félix, R., et al. Roles of endogenous ether lipids and associated PUFAs in the regulation of ion channels and their relevance for disease. J. Lipid Res. 61(6), 840-858 (2020).
Honsho, M. and Fujiki, Y. Plasmalogen homeostasis - regulation of plasmalogen biosynthesis and its physiological consequence in mammals. FEBS Lett. 591(18), 2720-2729 (2017).
Jenkins, C.M., Yang, K., Liu, G., et al. Cytochrome c is an oxidative stress-activated plasmalogenase that cleaves plasmenylcholine and plasmenylethanolamine at the sn-1 vinyl ether linkage. J. Biol. Chem. 293(22), 8693-8709 (2018).
Jiménez-Rojo, N. and Riezman, H. On the road to unraveling the molecular functions of ether lipids. FEBS Lett. 593(17), 2378-2389 (2019).
Mawatari, S., Sasuga, Y., Morisaki, T., et al. Identification of plasmalogens in Bifidobacterium longum, but not in Bifidobacterium animalis. Sci. Rep.10(1), 427 (2020).
Messias, M.C.F., Mecatti, G.C., Priolli, D.G., et al. Plasmalogen lipids: Functional mechanism and their involvement in gastrointestinal cancer. Lipids Health Dis.17(1), 41 (2018).
Paul, S., Lancaster, G.I., and Meikle, P.J. Plasmalogens: A potential therapeutic target for neurodegenerative and cardiometabolic disease. Prog. Lipid Res.74, 186-195 (2019).
Su, X.Q., Wang, J., and Sinclair, A.J. Plasmalogens and Alzheimer's disease: A review. Lipids Health Dis. 18(1), 100 (2019).
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640