News & Announcements
Essential Role of Esterified ω-Hydroxy Ceramides as Skin Lipids
Article from 2017-10-02
This article was originally published in the October 2017 edition of Matreya’s Newsletter for Glyco/Sphingolipid Research (PDF).
Ceramides in human cells have important and divergent functions that make their study both challenging and important. The roles of these ubiquitous lipids include signal transduction and cell regulation relevant to apoptosis, cell growth arrest, differentiation, senescence, and immune responses. Many of the functions of individual ceramides are dependent on the specific structure of each ceramide species. Relative to other tissues, human stratum corneum contains several very complex ceramide species that play important physicochemical roles in determining the cutaneous barrier's water-retaining functions.
The stratum corneum is the outermost cellular layer of the epidermis and functions as the permeability barrier in mammals. Of all the species of ceramides, the stratum corneum contains 12 extractable ceramide sub-groups including sphingosine, 6-hydroxysphingosine, dihydrosphingosine, and phytosphingosine bases. Mammalian skin contains significant amounts of sphingolipids (as much as 50% of the total lipids), particularly very long-chain linoleoyl esterified ceramide and glucosylceramide (also called O-acylceramide and O-acylglucosylceramide, respectively). These lipids, which are mostly found in the extracellular domains, are vital to the water permeability barrier’s ability to prevent lethal loss of water and pathogen invasion. The esterified ω-hydroxy ceramides are formed from glucosylceramide and sphingomyelin in special lamellar bodies in epidermal cells from which they are excreted into the stratum corneum. These skin-specific ceramides can be covalently bound to proteins of the cornified envelope where they form a hydrophobic layer.
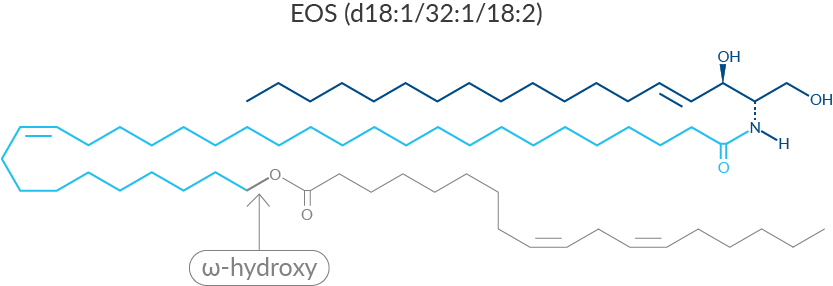
EOS (d18:1/32:1/18:2) is representative of the class of ceramides found in the outer layer of the epidermis.
A deficiency of linoleoyl ω-esterified ceramides is strongly correlated with skin diseases such as ichthyosis, psoriasis, and atopic dermatitis.1-3 Mutations in patatin-like phospholipase domain-containing 1 (PNPLA1), 12-lipoxygenase (12-LO), epidermal LO, and others, which play crucial roles in the biosynthesis of esterified ω-hydroxy ceramides by esterifying the ω-fatty acids with linoleic acid, have been shown to cause autosomal recessive congenital ichthyosis.4,5 A lack of activity in the enzyme serine palmitoyltransferase prevents the de novo synthesis of ceramides and has been strongly linked to the development of psoriasis.6 In atopic dermatitis, a deficiency of activity of the ELOVL (elongase of very long-chain fatty acids) enzymes results in a reduction in very long-chain fatty acids and a subsequent shortage of esterified long-chain ceramides.6
Due to the complex nature of the human stratum corneum, which contains dozens of ceramide species, analysis can be highly challenging. Shotgun lipidomics is a recent liquid chromatography-mass spectrometry technique that provides data on large amounts of analytes in a single experimental run. This approach allows for the rapid, high-throughput analysis of difficult samples. By taking advantage of specific esterified ω-hydroxy ceramide internal standards, whole classes of skin lipids have been identified and quantified.7
Available Skin Sphingolipids and Internal Standards
C18 Ceramide-d3 (d18:1/18:0-d3)
C18 dihydro Ceramide (d18:0/18:0)
C18 dihydro Ceramide-d3 (d18:0/18:0-d3)
C16 Sphingomyelin (d18:1/16:0)
C16 Sphingomyelin-13C (d18:1/16:0-13C)
C18 Ceramide-1-Phosphate-d3 (d18:1/18:0-d3)
C24 dihydro Ceramide (d18:0/24:0)
C30(ω-hydroxy) Ceramide (d18:1/30:0)
Read More about EOS in the Skin’s Lipid Matrix
Lipidomics Services
Our lipid experts have decades of experience in lipid synthesis, purification, and characterization with access to state-of-the-art equipment for discovery and targeted lipidomics.
Meet the Expert: Lipidomic Profiling Services
References
1. Breiden, B. and Sandhoff, K. The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochim. Biophys. Acta1841(3), 441-452 (2014).
2. Sandhoff, R. Very long chain sphingolipids: Tissue expression, function and synthesis. FEBS Lett. 584(9), 1907-1913 (2010).
3. Masukawa, Y., Narita, H., Shimizu, E., et al. Characterization of overall ceramide species in human stratum corneum. J. Lipid Res.49(7), 1466-1476 (2008).
4. Muñoz-Garcia, A., Thomas, C.P., Keeney, D.S., et al. The importance of the lipoxygenase-hepoxilin pathway in the mammalian epidermal barrier. Biochim. Biophys. Acta1841(3), 401-408 (2014).
5. Grond, S., Eichmann, T.O., Dubrac, S., et al. PNPLA1 deficiency in mice and humans leads to a defect in the synthesis of omega-O-acylceramides. J. Invest. Dermatol.137(2), 394-402 (2017).
6. Borodzicz, S., Rudnicka, L., Mirowska-Guzel, D., et al. The role of epidermal sphingolipids in dermatologic diseases. Lipids Health Dis. 15, 13 (2016).
7. Sadowski, T., Klose, C., Gerl, M.J. et al. Large-scale human skin lipidomics by quantitative, high-throughput shotgun mass spectrometry. Sci. Rep.7, 43761 (2017).
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640