News & Announcements
Advancing MASLD & MASH Research: Targeting De Novo Lipogenesis & Insulin Resistance
Article from 2024-08-28
Previous Post: Advancing MASLD & MASH Research: Tools & Strategies for Metabolic Dysfunction in Steatotic Liver Diseases
Understanding the mechanisms underlying de novo lipogenesis and its relationship to insulin resistance is important for studying metabolic associated steatotic liver disease (MASLD) and metabolic dysfunction-associated steatohepatitis (MASH), and, accordingly, developing effective treatments.
MASLD Therapeutic Strategies
Targeting De Novo Lipogenesis with Pharmacological Inhibitors
De novo lipogenesis contributes to hepatic lipid accumulation in MASLD through the formation of new fatty acids.1 New fatty acids are formed from non-lipid energy sources such as glucose, which is elevated by insulin resistance.2 Glucose is converted to acetyl-CoA, which is elongated into fatty acids via the fatty acid synthase (FASN) pathway. These fatty acids can be esterified to glycerol to produce triglycerides (TGs), which are stored in lipid droplets in the liver, leading to hepatic steatosis.1 Accordingly, strategies that decrease TG synthesis by targeting de novo lipogenesis reduce hepatic steatosis.
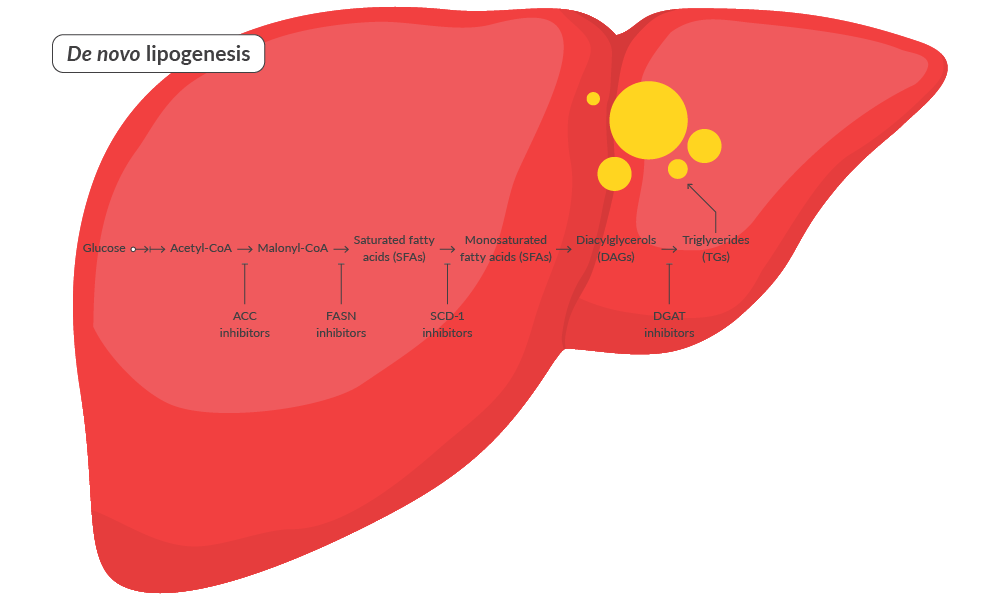
Strategies to Target De Novo Lipogenesis | ||
| Class | Effects | References |
| Acetyl-CoA carboxylase (ACC) inhibitors ACC inhibition suppresses the carboxylation of acetyl-CoA to malonyl-CoA, blocking fatty acid synthesis. | ↓ Fatty acid synthesis ↑ Fatty acid oxidation ↓ Hepatic triglyceride accumulation | 3 |
| Fatty acid synthase (FASN) inhibitors FASN inhibition blocks the synthesis of long-chain fatty acids from acetyl-CoA and malonyl-CoA to suppress fatty acid synthesis | ↓ Fatty acid synthesis ↓ Hepatic steatosis ↓ Inflammation | 4 |
| Stearoyl-CoA desaturase-1 (SCD-1) inhibitors SCD-1 inhibition suppresses the conversion of saturated fatty acids to monounsaturated fatty acids | ↓ Hepatic lipid accumulation ↑ Insulin sensitivity ↓ Inflammation | 5 |
| Diacylglycerol acyltransferase (DGAT) inhibitors DGAT inhibitors block the final step in triglyceride synthesis, reducing the formation of lipid droplets | ↓ Hepatic triglyceride synthesis ↓ Hepatic lipid accumulation | 6 |
De Novo Lipogenesis Assay Kits
| Item No. | Product Name | Description |
| 10010303 | Triglyceride Colorimetric Assay Kit | Measure TG levels in plasma, serum, cell lysates, and tissue homogenates |
| 500001 | Lipid Droplets Fluorescence Assay Kit | Detect lipid droplets, cellular organelles that are also referred to as lipid bodies, oil bodies, or adiposomes |
Targeting Insulin Resistance in the Development of MASLD
Insulin resistance plays a central role in the development of MASLD. Because of the excessive availability of glucose, de novo lipogenesis is increased by insulin resistance. In an insulin-resistant state, the inhibitory action of insulin on gluconeogenesis is suppressed, which increases gluconeogenesis, resulting in increased glucose production and availability for de novo lipogenesis, exacerbating hepatic lipid accumulation.7 It also blocks the inhibition of lipolysis in adipose tissue, leading to the release of free fatty acids (FFAs), which are transported to the liver, further contributing to hepatic lipid accumulation.8 Accordingly, therapies designed for the treatment of type 2 diabetes have been repurposed for the treatment of MASLD.
Strategies to Target Insulin Resistance | ||
| Class | Effects | References |
| GLP-1R agonists Activation of glucagon-like peptide-1 receptor (GLP-1R) mimics the actions of GLP-1 to regulate glucose homeostasis | ↑ Glucose-stimulated insulin secretion ↑ Insulin sensitivity ↓ Blood glucose levels ↓ Hepatic steatosis | 7,9 |
| SGLT2 inhibitors Blocking the activity of sodium-glucose cotransporter 2 (SGLT2) in the kidneys decreases glucose reabsorption | ↓ Renal glucose reabsorption ↑ Glucose excretion ↓ Blood glucose levels ↓ Hepatic steatosis | 7,9-11 |
| DPP-4 inhibitors Inhibiting the activity of DPP-4, an enzyme that degrades incretin hormones like GLP-1 and gastric inhibitory peptide (GIP), increases the activity of these hormones | ↑ GLP-1 and GIP activity ↑ Glucose-stimulated insulin secretion ↓ Blood glucose levels | 7,12,13 |
Insulin & Glucose Homeostasis Assay Kits
| Item No. | Product Name | Description |
| 26619 | Insulin (human) ELISA Kit | For the measurement of insulin in cell culture supernatant, serum, and plasma |
| 589501 | Insulin (mouse, rat) EIA Kit | For the measurement of insulin in rat, human, mouse, and other samples |
| 500305 | THUNDER™ Human Insulin TR-FRET Biomarker Assay Kit | A sandwich TR-FRET immunoassay for the quantitative measurement of human insulin in cell culture supernatants |
| 10009582 | Glucose Colorimetric Assay Kit | For analysis of glucose in plasma, serum, and urine |
| 600470 | Glucose Uptake Cell-Based Assay Kit | A cell-based assay for studying modulators of cellular glucose uptake |
Strategies Targeting Excess FFAs
Under normal conditions, insulin inhibits lipase, decreasing the liberation of FFAs.8 However, in an insulin-resistant state, this inhibitory action is blocked, allowing lipase to liberate FFAs from TGs stored in adipose tissue, which increases the flux of FFAs transported to the liver. In the liver, these FFAs are converted to TGs, contributing to lipid droplet formation and hepatic steatosis.
Accordingly, lipase inhibitors block the release of FFAs, reducing the pool of available substrates for TG synthesis.
Strategies to Target FFA Release | ||
Class | Effects | References |
| Lipase inhibitors By inhibiting TG hydrolysis in adipose tissue, FFA release and subsequent transport to the liver is blocked | ↓ FFA levels | 8,14 |
FFA Assay Kits
| Item No. | Product Name | Description |
| 700310 | Free Fatty Acid Fluorometric Assay Kit | Measure FFAs in plasma, serum, and urine |
| 700640 | Lipase Activity Assay Kit | A fluorescence-based method for detecting lipase activity in plasma, serum, tissue homogenates, and cell culture samples |
Stimulation of Fatty Acid Oxidation to Decrease FFAs
The main pathway for the degradation of FFAs is mitochondrial β-oxidation.15 However, sustained mitochondrial β-oxidation leads to increased levels of reactive oxygen species (ROS), which induce inflammation and cellular injury, and eventual mitochondrial dysfunction, which impairs β-oxidation.16 Impaired mitochondrial function can be overcome by enhancing fatty acid oxidation through activators of this pathway.
Strategies to Target Fatty Acid Oxidation | ||
| Class | Effects | References |
| AMPK activators AMPK activation increases ATP production and decreases ATP consumption to improve energy balance | ↑ Hepatic fatty acid oxidation ↓ De novo lipogenesis ↑ Mitochondrial function | 15,17,18 |
Mitochondrial Function Assay Kits
| | MitoCheck® AssaysMeasure the activity of complexes I-V of the electron transport chain with Cayman's MitoCheck® Assays. |
Cellular Metabolism Analysis
Explore mitochondrial function dysregulation using high-content imaging, enzymatic screening, and the Agilent Seahorse™ XF Pro.
|
Oxidative Stress Assay Kits & Fluorescent Probes
| | Cayman offers a collection of assays for both the direct measurement of reactive species as well as markers of oxidative stress. Download the Oxidative Stress: Markers & Detection Tools Guide |
The interplay between de novo lipogenesis and insulin resistance contributes significantly to hepatic steatosis, a hallmark of MASLD. However, given the complexity and heterogeneity of MASLD and MASH, other approaches to regulating lipid homeostasis should also be considered. One such approach is by targeting fatty acid signaling pathways, including peroxisome proliferator-activated receptors (PPARs) and free fatty acid receptors (FFARs), a topic covered in the next post.
Next Post: Advancing MASLD & MASH Research: PPAR and FFAR Fatty Acid Signaling Pathways
Related Posts:
- Advancing MASLD & MASH Research: An Introduction
- Advancing MASLD & MASH Research: The Roles of LXRs, FXRs, and GP-BAR1 in Metabolic Regulation
Products for MASLD & MASH Research
The products featured in this series are a snapshot of the comprehensive resources available from Cayman for MASLD and MASH research. Our full catalog contains a comprehensive range of biochemicals, proteins, antibodies, and assay kits to support MASLD and MASH research.
View all MASLD-related products
References
1. Esler, W.P. and Cohen, D.E. Pharmacologic inhibition of lipogenesis for the treatment of NAFLD. J. Hepatol. 80(2), 362-377 (2024).
2. Batchuluun, B., Pinkosky, S.L., and Steinberg, G.R. Lipogenesis inhibitors: Therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 21(4), 283-305 (2022).
3. Chen, L., Duan, Y., Huiqiang, W., et al. Acetyl-CoA carboxylase (ACC) as a therapeutic target for metabolic syndrome and recent developments in ACC1/2 inhibitors. Expert Opin. Investig. Drugs 28(10), 917-930 (2019).
4. Teng, T., Qiu, S., Zhao, Y., et al. Pathogenesis and therapeutic strategies related to non-alcoholic fatty liver disease. Int. J. Mol. Sci. 23(14), 7841 (2022).
5. Jeyakumar, S.M. and Vajreswari, A. Stearoyl-CoA desaturase 1: A potential target for non-alcoholic fatty liver disease?-perspective on emerging experimental evidence. World J. Hepatol. 14(1), 168-179 (2022).
6. Sui, X., Wang, K., Song, K., et al. Mechanism of action for small-molecule inhibitors of triacylglycerol synthesis. Nat. Commun. 14(1), 3100 (2023).
7. Shao, Y., Chen, S., Han, L. et al. Pharmacotherapies of NAFLD: Updated opportunities based on metabolic intervention. Nutr. Metab. (Lond) 20(1), 30 (2023).
8. Pal, S.C. and Méndez-Sánchez, N. Insulin resistance and adipose tissue interactions as the cornerstone of metabolic (dysfunction)-associated fatty liver disease pathogenesis. World J. Gastroenterol. 29(25), 3999-4008 (2023).
9. Nevola, R., Epifani, R., Imbriani, S., et al. GLP-1 receptor agonists in non-alcoholic fatty liver disease: Current evidence and future perspectives. Int. J. Mol. Sci. 24(2), 1703 (2023).
10. Fonseca-Correa, J.I. and Correa-Rotter, R. Sodium-glucose cotransporter 2 inhibitors mechanisms of action: A review. Front. Med. (Lausanne) 8, 777861 (2021).
11. Raj, H., Durgia, H., Palui, R., et al. SGLT-2 inhibitors in non-alcoholic fatty liver disease patients with type 2 diabetes mellitus: A systematic review. World J. Diabetes 10(2), 114-132 (2019).
12. Makrilakis, K. The role of DPP-4 inhibitors in the treatment algorithm of type 2 diabetes mellitus: When to select, what to expect. Int. J. Environ. Res. Public Health 16(15), 2720 (2019).
13. Gilbert, M.P. and Pratley, R.E. GLP-1 analogs and DPP-4 inhibitors in type 2 diabetes therapy: Review of head-to-head clinical trials. Front. Endocrinol. (Lausanne) 11, 178 (2020).
14. Grabner, G.F., Xie, H., Schweiger, M. et al. Lipolysis: Cellular mechanisms for lipid mobilization from fat stores. Nat. Metab. 3(11), 1445-1465 (2021).
15. Houten, S.M., Violante, S., Ventura, F.V., et al. The biochemistry and physiology of mitochondrial fatty acid β-oxidation and its genetic disorders. Annu. Rev. Physiol. 78, 23-44 (2016).
16. Arumugam, M.K., Gopal, T., Kandy, R.R.K., et al. Mitochondrial dysfunction-associated mechanisms in the development of chronic liver diseases. Biology (Basel) 12(10), 1311 (2023).
17. Herzig, S. and Shaw, R. J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 19(2), 121-135 (2018).
18. Fang, C., Pan, J., Qu, N., et al. The AMPK pathway in fatty liver disease. Front. Physiol. 13, 970292 (2022).
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640