Technical Support & Resources
Information provided in the product description is from published literature. Due to the nature of scientific experimentation, your results (e.g., selectivity and effective concentrations) or specific application for this product may differ. If you have questions about how this product fits your application, please contact our technical support staff.
Visit our FAQ
Contact Us
Toll Free Phone (USA and Canada Only): (888) 526-5351
Direct Phone: (734) 975-3888
Product Categories
Application

Technical Information
- Bovine Serum Albumin-PA
- Bovine Serum Albumin-Palmitate
- BSA-Hexadecanoic Acid
- BSA-PA
Shipping & Storage Information
Recommended Products
Certificates of Analysis & Batch Specific Data
Provide batch numbers separated by commas to download or request available product inserts, QC sheets, certificates of analysis, data packs, and GC-MS data.
Lipid Resource Center
Discover Products & Resources for Lipid Research
- High-purity lipid standards
- Lipid roles in biology
- Lipids in health & disease
- Lipids for pharmaceutical development
- Protocols, advice, & resources
Product Description
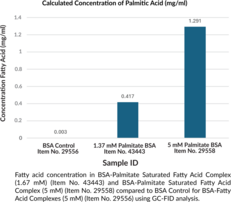
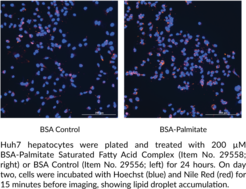
BSA-Palmitate Saturated Fatty Acid Complex (5 mM) is composed of palmitic acid (Item No. 10006627) and fatty acid-free bovine serum albumin (BSA) at an approximately 6:1 molar ratio of palmitate:BSA. It was prepared under sterile conditions, then filtered and aliquoted into sterile vials. Cayman’s BSA-Palmitate Saturated Fatty Acid Complex (5 mM) can be used for efficient fatty acid delivery to cells in culture for the purpose of monitoring lipid metabolism, including fatty acid oxidation, and inflammatory signaling pathways, such as in studies of obesity-related inflammation and insulin resistance.1,2,3 It has been used to study the effect of long-chain fatty acid uptake on gene and protein expression, lipid droplet formation, hepatocyte lipid accumulation, and oxidative stress.4,5,6 Cayman’s BSA-Palmitate Saturated Fatty Acid Complex (5 mM) is suitable for use in short- and long-term cell culture applications (25+ hours). For best results, it is recommended that this product be used in conjunction with Cayman’s BSA Control for BSA-Fatty Acid Complexes (5 mM) (Item No. 29556), prepared with fatty acid-free BSA. A BSA-palmitate complex is also available as part of Cayman's BSA-Palmitate Reagent Set (5 mM) (Item No. 44698).
WARNING This product is not for human or veterinary use.
References & Product Citations
1. Cell culture models of fatty acid overload: Problems and solutions. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1863(2), 143-151 (2018).
2. Oxygen flux analysis to understand the biological function of sirtuins. Methods Mol. Biol. 1077, 241-258 (2013).
3. Inhibition or deletion of the lipopolysaccharide receptor Toll-
4. Deletion of PGAM5 downregulates FABP1 and attenuates long-
5. Inhibition of hepatic oxalate overproduction ameliorates metabolic dysfunction-
6. Short-
Epstein-
MicroRNA 29a alleviates mitochondrial stress in diet-
Increased fatty acid delivery by tumor endothelium promotes metastatic outgrowth. JCI Insight 10(9), e187531 (2025).
RhoA regulates membrane order and tension to control excitability of nociceptor neurons. bioRxiv [Preprint] (2025).
Glutamate decreases oxidative stress and lipid droplet formation in astrocytes. J. Cell Sci. 138(19), jcs263983 (2025).
CNS-
Regional metabolic analysis of structurally preserved kidney slices by ex vivo respirometry. Am. J. Physiol. Renal Physiol. 329(6), F796-F808 (2025).
Altered fatty acid oxidation via CPT1A promotes epithelial-
Interaction of sortilin with apolipoprotein E3 enables neurons to use long-
Enhanced nuclear localization of small heterodimer partner in metabolic dysfunction-
A retinoic acid: YAP1 signaling axis controls atrial lineage commitment. Cell Rep. 44(5), 115687 (2025).
Fatty acid desaturases link cell metabolism pathways to promote proliferation of Epstein-
Perturbation of de novo lipogenesis hinders MERS-
Membrane lipid nanodomains modulate HCN pacemaker channels in nociceptor DRG neurons. Nat. Commun. 15(1), 9898 (2024).
Reducing the mitochondrial oxidative burden alleviates lipid-
A metabolic dysfunction-
An intrinsic mechanism of metabolic tuning promotes cardiac resilience to stress. EMBO Mol. Med. 16(10), 2450-2484 (2024).
Inhibition of hepatic oxalate overproduction ameliorates metabolic dysfunction-
Transcriptomic, epigenomic, and spatial metabolomic cell profiling redefines regional human kidney anatomy. Cell Metab. 36(5), 1105-1125 (2024).
Endogenous renal adiponectin drives gluconeogenesis through enhancing pyruvate and fatty acid utilization. Nat. Commun. 14(1), 6531 (2023).
Deletion of PGAM5 downregulates FABP1 and attenuates long-
Obesity-
Brown adipose tissue-
The dynamic clustering of insulin receptor underlies its signaling and is disrupted in insulin resistance. Nat. Commun. 13(1), 7522 (2022).
Comprehensive single-
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640