News & Announcements
Vaccine Development for Emerging Infectious Disease
Article from 2020-04-13
STING Adjuvants in Viral and Bacterial Vaccines
By Melissa A. Bates, Ph.D. -Technical Writer, Cayman Chemical
Vaccines Prevent Widespread Contagious Diseases
The pandemic caused by the unexpected emergence of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the causative agent of COVID-19, has prompted an urgent need to develop a vaccine against this infectious disease. Vaccines provide protection against infectious diseases by inducing immunological memory to a pathogen.
The Immunology of Vaccines
Immunological memory is a hallmark of the immune system that confers long-term protection against infectious agents. This property can be exploited through vaccines. Vaccines are formulated to initiate an innate immune response that directs the antigen-specific adaptive immune response. Pathogen-associated molecular patterns (PAMPs) are pathogen-specific signatures that are recognized by cells of the innate immune system, including macrophages, neutrophils, and dendritic cells (DCs), through a diverse repertoire of pattern recognition receptors (PRRs). PRRs that sense viral PAMPs include the endosomal toll-like receptors (TLRs), TLR3, TLR7, TLR8, and TLR9, as well as cytosolic nucleic acid sensors, including nucleotide-binding oligomerization domain-containing protein 1 (NOD1) , NOD2, retinoic acid-inducible gene I (RIG-I), melanoma differentiation-associated protein 5 (MDA5), and cyclic GMP-AMP (cGAMP) synthase (cGAS). Adjuvants are substances included in a vaccine that boost the antigen-specific immune response by activating PRRs. They activate PRRs by either directly acting as PRR ligands themselves or by inducing the release of damage-associated molecular patterns (DAMPs), such as uric acid, that activate PRRs. Many various PRR agonists are available from Cayman.
Available PRR Agonists
Pam3CSK4 (trifluoroacetate salt)
Toll-Like Receptor 7 Ligand II
TLR7/8 Agonist 1 (hydrochloride)
Activation of PRRs that have a signature immunostimulatory profile tailors the adaptive immune response towards one that is most effective against the target pathogen. It induces signal transduction pathways that result in a distinct profile of gene and co-stimulatory molecule expression, as well as the release of cytokines, chemokines, and other immunomodulators that direct the adaptive immune response. After antigen encounter at the site of vaccination, DCs migrate to the lymph nodes and present antigens in surface major histocompatibility complexes (MHCs) to antigen-specific T cells, which differentiate into T helper cells. Different subsets of T helper cells can influence the nature of the resulting immune responses to optimally eradicate the perceived threat. After the initial immune response, a small number of antigen-specific cells differentiate into long-lived, memory T and B cells. Upon re-exposure to the antigen, memory T and B cells rapidly expand into effector T cells and plasma cells, respectively, enabling a rapid response to the pathogen and preventing infectious diseases.
Vaccines provide defense against bacterial and viral pathogens by inducing immunological memory.
Type I Interferons Are Critical for Antiviral Immunity
Type I interferons (IFN-α and IFN-β) are critical for host defense against viral pathogens. Type I IFNs induce the transcription of a variety of interferon-stimulated genes that act in an autocrine, paracrine, or systemic manner to induce a myriad of effects on the immune system that collectively facilitate defense against viral pathogens. They reduce viral replication in infected cells and induce an antiviral state in neighboring cells. Antigen-presenting cells (APCs) stimulated with type I IFNs increase surface expression of MHC and co-stimulatory molecules, increasing the ability of APCs to stimulate differentiation of naïve T cells into effector T cells. Type I IFNs also promote the induction and proliferation of memory T cells.
STING Activation by Vaccine Adjuvants
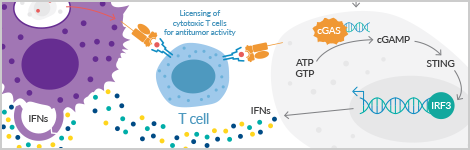
Ligands that activate STimulator of INterferon Genes (STING) induce type I IFNs and have been used as vaccine adjuvants in preclinical models. STING is a signaling protein located on the endoplasmic reticulum and is widely expressed in immune cells. The cyclic dinucleotides (CDNs) cyclic di-GMP, cyclic di-AMP, and 3’3’-cGAMP are bacterial second messengers that bind to and activate STING. STING can also be activated through the action of cGAS. cGAS is a cytosolic DNA sensor that catalyzes the formation of the STING activator 2’3’-cGAMP upon recognition of foreign DNA, including viral DNA. Cayman, in conjunction with the expert nucleotide scientists at Biolog Life Science Institute in Germany, has developed cyclic di-GMP ELISA, cyclic di-AMP ELISA, 3'3'-cGAMP ELISA, and 2'3'-cGAMP ELISA kits for specific detection of these CDNs in bacterial and mammalian cell lysates.
STING activation recruits the adapter protein TANK-binding kinase 1 (TBK1), which phosphorylates and activates the transcription factor interferon regulatory factor 3 (IRF3), leading to the expression of type I IFNs. STING also activates NF-ĸB, resulting in the production of additional inflammatory cytokines. STING has also been shown to interact with RIG-I and mitochondrial antiviral-signaling protein (MAVS), which are key cytosolic sensors of viral RNA, though the precise molecular events leading to STING activation have yet to be elucidated.
STING activators induce IFN and pro-inflammatory cytokines.
Natural STING ligands have been used to boost vaccine efficacy against viral and bacterial pathogens. Wang et al. demonstrated that 2’3’-cGAMP preferentially induced a Th1-mediated immune response that was associated with improved survival upon viral challenge when used as an adjuvant in the H1N1 swine influenza vaccine in mice. These authors also found that mice administered an H5N1 avian influenza vaccine containing 2’3’-cGAMP as an adjuvant developed antigen-specific antibodies within two weeks of immunization that remained in circulation for at least 40 weeks. Blaauboer et al. showed that intranasal administration of cyclic di-GMP increases ovalbumin (OVA) uptake and processing by pulmonary DCs and decreases lung colony forming units in a mouse model of lung S. pneumoniae infection.
STING ligands have also shown promise as cancer vaccine adjuvants. Gutjahr et al. demonstrated that immunization with OVA and 2’3’-cGAMP increased the percentage of OVA-specific IFN-γ+ CD8+ T cells and inhibited tumor growth in an OVA+ EG7 tumor implant mouse model. To measure the efficacy of OVA/adjuvant immunization in mice, Cayman offers ELISAs to quantitate anti-OVA IgG1 and IgE.
Nucleoside- and non-nucleoside-based STING activators have been successfully used as vaccine adjuvants in preclinical models. STING activators available from Cayman can be found below. Inhibition of ectonucleotide pyrophosphatase/phosphodiesterase 1 (ENPP1), a cGAMP hydrolase, is an additional approach that could be investigated for use in vaccines. Inhibitors of ENPP1 increase the concentration of 2’3’-cGAMP by reducing its degradation and thus could be further explored as a means to increase STING activation.
Key Considerations for Vaccine Development
Successful implementation of a vaccine-based population-wide infectious disease control strategy requires that the vaccine is safe and effective in all individuals, including those most susceptible to infectious diseases, such as children, the elderly, and immunocompromised individuals.
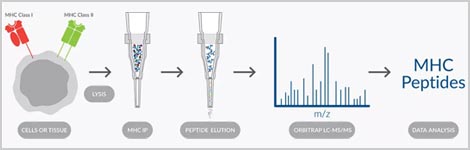
Antigens used in vaccines must be selectively and stably expressed by the pathogen. It should distinguish between commensal and pathogenic bacterial or viral strains and not subject to geographic or temporal variability. Cayman offers customizable Immunopeptidome Profiling Services to assist in the identification of neoantigens for vaccines. Our scientists sequence antigens presented by MHC and identify those unique to the pathogen. Using this approach, it is possible to not only identify novel antigens, but to determine individual differences in antigen presentation that could be used to develop personalized vaccines that capture the diversity of the human population. The combination of antigen and adjuvant used in a vaccine must be balanced to have sufficient immunogenicity without overt reactogenicity. Cayman has human and mouse multiplex kits to simultaneously measure multiple cytokines in a single sample in a convenient and cost-effective platform.
Vaccines are a preventative approach to infectious diseases that requires vigilance for emerging pathogens. However, the utility of a vaccination program to control an ongoing pandemic caused by a novel pathogen is hindered by the time required for their design and is inevitably reliant on the availability of existing agents to inhibit pathogen replication and manage symptoms of the infectious disease. Cayman has assembled an Antiviral Screening Library containing more than 410 known antiviral compounds that could identify agents that can be used to research novel pandemic viruses for which there are no vaccines or existing therapeutics, such as SARS-CoV-2. Complementary to this are SARS-CoV-2 detection assays that can be used to identify SARS-CoV-2 infection in research samples.
Learn more about
Cyclic Dinucleotides - Ubiquitous Cellular Messengers
Available STING Activators
diABZI STING Agonist 3 (hydrochloride)
You May Also Be Interested In
 |  |
|

References
Bastola, R., Noh, G., Keum, T. et al. Vaccine adjuvants: Smart components to boost the immune system. Arch. Pharm. Res.40(11), 1238-1248 (2017).
Blaauboer, S.M., Mansouri, S., Tucker, H.R. et al. The mucosal adjuvant cyclic di-GMP enhances antigen uptake and selectively activates pinocytosis-efficient cells in vivo. Elife 4, e06670 (2015).
Dubensky, T.W., Jr., Kanne, D.B., and Leong, M.L. Rationale, progress, and development of vaccines utilizing STING-activating cyclic dinucleotide adjuvants. Ther. Adv. Vaccines 1(4), 131-143 (2013).
Ni, G., Ma, Z., and Damania, B. cGAS and STING: At the intersection of DNA and RNA virus-sensing networks. PLoS Pathog. 14(8), e1007148 (2018).
Gutjahr, A., Papagno, L., Nicoli, F. et al. The STING ligand cGAMP potentiates the efficacy of vaccine-induced CD8+ T cells. JCI Insight 4(7), e125107 (2019).
Huber, J.P. and Farrar, J.D. Regulation of effector and memory T-cell functions by type I interferon. Immunology 132(4), 466-474 (2011).
Shin, J.-H., Lee, J.-H., Jeong, S.D. et al. C-di-GMP with influenza vaccine showed enhanced and shifted immune responses in microneedle vaccination in the skin. Drug Deliv. Transl. Res. 10(3), 815-825 (2020).
Stern, P.L. Key steps in vaccine development. Ann. Allergy Asthma Immunol. 125(1), 17-27 (2020).
Su, T., Zhang, Y, Valerie, K. et al. STING activation in cancer immunotherapy. Theranostics 9(25), 7759-7771 (2019).
Volckmar J., Knop, L, Stegemann-Koniszewski, S. et al. The STING activator c-di-AMP exerts superior adjuvant properties than the formulation poly(I:C)/CpG after subcutaneous vaccination with soluble protein antigen or DEC-205-mediated antigen targeting to dendritic cells. Vaccine 37(35), 4963-4974 (2019).
Wang, J., Li, P., and Wu, M.X. Natural STING agonist as an “ideal” adjuvant for cutaneous vaccination. J. Invest. Dermatol.136(11), 2183-2191 (2016).
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640