News & Announcements
Bacterial Cyclic Dinucleotides Regulate Biofilm Formation
Article from 2022-09-16
Many species of bacteria possess the ability to adhere to biological or synthetic surfaces and form multicellular communities known as biofilms. An estimated 65-80% of bacterial infections are associated with biofilms, including chronic wound infections, endocarditis, otitis media, medical implant contamination, and chronic lung infections in patients with cystic fibrosis.1,2 Major human pathogens known to cause biofilm-associated disease include Escherichia coli, Pseudomonas aeruginosa, Enterococcus faecalis, Staphylococcus aureus, and Streptococcus pneumoniae.
Within a biofilm, bacteria are encased in an extracellular matrix comprised of exopolysaccharides (EPS), proteins, lipids, and nucleic acids that protects them from clearance by the host immune system or antibiotics (Figure 1). In fact, bacteria are 1,000 times more tolerant to antibiotics when they are encased in a biofilm compared with existing in a motile, planktonic state.1 Finding new ways to disrupt biofilm formation or persistence may help facilitate treatment of biofilm-associated infections.
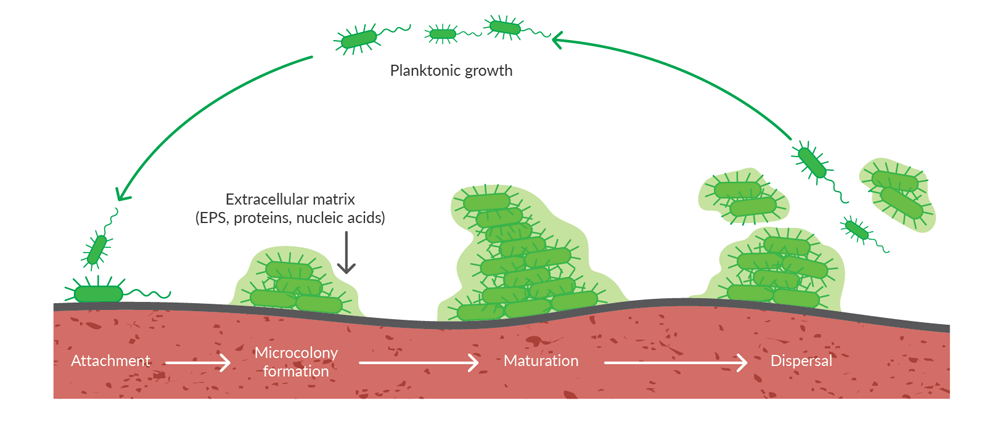
Figure 1. Bacterial biofilm formation and dispersal. Following attachment of motile, planktonic bacteria to a surface, cell division and production of extracellular matrix components leads to formation of microcolonies that mature into complex biofilms. Individual cells can disperse from the biofilm to resume a planktonic lifestyle or sections of the biofilm can be dispersed by mechanical disruption.
Many bacteria regulate the conversion between planktonic and biofilm lifestyles through cyclic dinucleotide (CDN) second messengers, including cyclic di-GMP (c-di-GMP), cyclic di-AMP (c-di-AMP), and 3'3'-cyclic GMP-AMP (3'3'-cGAMP). Modulation of CDN synthesis and signaling may provide new avenues for treatment of biofilm-associated infections.
Sensitive, Accurate Quantification of CDNs
Cayman, in conjunction with the expert nucleotide scientists at Biolog Life Science Institute in Germany, has developed immunoassays for specific detection of 3’3’-cGAMP, cyclic di-GMP, and cyclic di-AMP in bacterial cell lysates using a colorimetric 96-well microtiter plate format. By monitoring CDN levels in biological samples, these assays can be used to identify compounds that modulate CDN synthesis, degradation, and signaling.
| 3’3’-cGAMP ELISA Kit | Cyclic di-GMP ELISA Kit | Cyclic di-AMP ELISA Kit |
|
|
|
c-di-GMP
c-di-GMP is a major activator of biofilm formation that signals through various proteins and riboswitches to regulate biofilm-related processes such as bacterial adhesion, motility, and EPS synthesis.3,4 c-di-GMP is synthesized from GTP by diguanylate cyclase (DGC) enzymes possessing a GGDEF domain and degraded by phosphodiesterases (PDEs) containing HD-GYP or EAL domains into 5'-pGpG or GMP, respectively (Figure 2).3 While environmental cues such as oxygen, light, and reducing conditions are known to effect c-di-GMP synthesis and turnover by DGCs and PDEs, much is still unknown about how these enzymes are regulated.4,5
Figure 2. Synthesis and degradation of c-di-GMP.
c-di-AMP
c-di-AMP is perhaps best known for its roles in regulating cellular osmolarity and potassium homeostasis, but has also been shown to influence biofilm formation.5,6 It regulates EPS synthesis in Streptococcus mutans and Bacillus subtilis, but has opposite effects on biofilm formation in these bacteria.7,8 c-di-AMP is synthesized from two molecules of ATP by enzymes with diadenylate cyclase (DAC) activity and is degraded into 5'-pApA or AMP by PDEs that have either a DHH-DHHA1 domain or an HD domain (Figure 3).5,7 Accumulation of c-di-AMP can be influenced by environmental factors including potassium availability or nitrogen source.6 Recently, it was demonstrated using Cayman's Cyclic di-AMP ELISA Kit that the bile salts cholate and deoxycholate decrease c-di-AMP levels in the enteropathogen Clostridioides difficile.9 Aside from regulating the activities of DAC and PDE enzymes, some bacteria such as Listeria monocytogenes can regulate intracellular c-di-AMP levels through the use of multidrug efflux pumps (MDRs) to export c-di-AMP from cells.8
Figure 3. Synthesis and degradation of c-di-AMP.
3'3'-cGAMP
3’3’-cGAMP has been shown to regulate processes including phage response, osmotic stress response, bacterial motility, and biofilm formation.5 In Vibrio cholerae, 3’3’-cGAMP is synthesized from ATP and GTP by DncV, which belongs to the cGAS/DncV-like nucleotidyltransferase (CD-NTase) superfamily (Figure 4). DncV is structurally analogous to the cyclic GMP-AMP synthase (cGAS) enzyme that synthesizes 2’3’-cGAMP in eukaryotes. In deltaproteobacteria, however, 3’3’-cGAMP is instead synthesized by Hypr GGDEF proteins. 3’3’-cGAMP is degraded by phosphodiesterases (PDEs) including PmxA in Myxococcus xanthus and V-cGAP1, V-cGAP2, V-cGAP3, and VcEAL in V. cholerae.
Figure 4. Synthesis and degradation of 3’3’-cGAMP.
Antibiofilm Strategies Target CDN Signaling
Most of the research into antibiofilm strategies targeting CDN signaling has focused on c-di-GMP due to its function as a major regulator of biofilm formation, the conservation of c-di-GMP signaling across many bacterial species, and a lack of c-di-GMP-associated enzymes in eukaryotes. Small molecule inhibitors of DGC enzymes that synthesize c-di-GMP have been shown to reduce biofilm formation. For example, DI-3 and DI-10 inhibit DGC enzymes found in V. cholerae, reduce intracellular levels of c-di-GMP, and prevent biofilm formation.1 The small molecule ebselen inhibits c-di-GMP binding to the DGC enzyme WspR as well as PelD, a c-di-GMP effector involved in EPS biosynthesis, in P. aeruginosa, where it increases swimming motility and reduces biofilm formation.2 Another small molecule inhibitor of WspR, LP 3134, inhibits biofilm formation by P. aeruginosa and Acinetobacter baumannii, reduces adherence of P. aeruginosa to silicone catheters, and disperses established biofilms of both of these bacteria.10
A similar strategy has also been used to target c-di-AMP signaling. DisA is a bacterial enzyme possessing c-di-AMP synthase activity and a DAC domain.11 ST056083 is a small molecule inhibitor of DisA that inhibits growth, EPS synthesis, and biofilm formation in E. faecalis, a bacterium that can cause urinary tract infections, endocarditis, and various oral diseases. Other inhibitors of DisA include the polyphenols tannic acid (gallotannin), theaflavin-3'-gallate, theaflavin 3,3'-digallate, and the antiparasitic drug suramin.2
Another approach to modulate c-di-GMP signaling includes sequestration or inactivation of this CDN using compounds such as auramine O, which forms c-di-GMP aggregates.2 PDE enzymes that degrade c-di-GMP also present attractive targets. In P. aeruginosa, for example, nitric oxide enhances PDE activity, which reduces intracellular levels of c-di-GMP and promotes the dispersal of biofilms.12
Given the therapeutic challenges of treating biofilm-associated infections, new antibiofilm strategies are needed to disrupt biofilms and make bacteria more susceptible to host immune responses and antimicrobial treatments. Given their roles in biofilm formation, CDN signaling pathways present attractive targets for new antibiofilm therapies. Cayman Chemical offers a wide range of products for the elucidation of these signaling pathways and their regulation, including assay kits, CDNs, and modulators of CDN biosynthesis.
Assay Kits
3'3'-cGAMP ELISA Kit
Cyclic di-GMP ELISA Kit
Cyclic di-AMP ELISA Kit)
CDNs, Metabolites, and Negative Controls
3'3'-cGAMP (sodium salt)
5'-pApA (sodium salt)
5'-pGpG (sodium salt)
Cyclic di-AMP (sodium salt)
Cyclic di-GMP (sodium salt)
Cyclic di-IMP (sodium salt)
Cyclic di-UMP (sodium salt)
Cayman also provides a variety of Services to support the investigation of CDN signaling pathways and development of novel antibiofilm agents, including protein and antibody production, activity screening and assay development, and chemical synthesis services.
You May Also Be Interested In



References
1. Sambanthamoorthy, K., Sloup, R.E., Parashar, V., et al. Identification of small molecules that antagonize diguanylate cyclase enzymes to inhibit biofilm formation. Antimicrob. Agents Chemother. 56(10), 5202-5211 (2012).
2. Martín-Rodríguez, A.J. and Römling, U. Nucleotide second messenger signaling as a target for the control of bacterial biofilm formation. Curr. Top. Med. Chem. 17, 1928-1944 (2017).
3. da Purificaҫão, A.D., de Azevedo, N.M., de Araujo, G.G., et al. The world of cyclic dinucleotides in bacterial behavior. Molecules25(10), 2462 (2020).
4. Römling, U. and Balsalobre, C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J. Intern. Med. 272(6), 541-561 (2012)
5. Yoon, S.H. and Waters, C.M. The ever-expanding world of bacterial cyclic oligonucleotide second messengers. Curr. Opin. Microbiol. 60, 96-103 (2021).
6. Stülke, J. and Krüger, L. Cyclic di-AMP signaling in bacteria. Annu. Rev. Microbiol. 74, 159-179 (2020).
7. Xiong, Z.-Q., Fan, Y.-Z., Song, X., et al. The second messenger c-di-AMP mediates bacterial exopolysaccharide biosynthesis: A review. Mol. Biol. Rep. 47(11), 9149-9157 (2020).
8. Yin, W., Cai, X., Ma, H., et al. A decade of research on the second messenger c-di-AMP. FEMS Microbiol. Rev.44(6), 701-724 (2020).
9. Oberkampf, M., Hamiot, A., Altamirano-Silva, P., et al. c-di-AMP signaling is required for bile salt resistance, osmotolerance, and long-term host colonization by Clostridioides difficile. Sci. Signal. 15(750), eabn8171 (2022).
10. Sambanthamoorthy, K., Luo, C., Pattabiraman, N., et al. Identification of small molecules inhibiting diguanylate cyclases to control bacterial biofilm development. Biofouling30(1), 17-28 (2014).
11. Chen, L., Li, X., Zhou, X., et al. Inhibition of Enterococcus faecalis growth and biofilm formation by molecule targeting cyclic di-AMP synthetase activity. J. Endod.44(9), 1381-1388 (2018).
12. Caly, D.L., Bellini, D., Walsh, M.A., et al. Targeting cyclic di-GMP signalling: A strategy to control biofilm formation? Curr. Pharm. Des. 21(1), 12-24 (2015).
Contact Info
Cayman Chemical1180 East Ellsworth RoadAnn Arbor, Michigan 48108 USAToll Free: (800) 364-9897(USA and Canada only)Fax: (734) 971-3640