Cancer Metabolism
Glucose Metabolism
Metabolic reprogramming is a hallmark of cancer cells that increases survival in nutrient-poor or otherwise stressful environments and promotes elevated levels of growth and proliferation. A classic example of metabolic reprogramming is the Warburg effect, or aerobic glycolysis, whereby cancer cells utilize glycolysis regardless of the presence of oxygen. These cells exhibit a high rate of glucose uptake and lactate production. This process is supported by upregulation of glucose transporters, including Glut1. Glycolysis generates ATP and NADH, as well as a variety of carbon-based intermediates that can branch off into biosynthetic pathways, including the pentose phosphate and serine biosynthesis pathways, to generate NADPH, amino acids, nucleotides, and fatty acids.
VIEW ALL CARBOHYDRATE METABOLISM PRODUCTSGlucose Metabolism
Explore our tools to study glucose metabolism, including assay kits, substrates, and inhibitors.
DOWNLOAD THE BROCHURE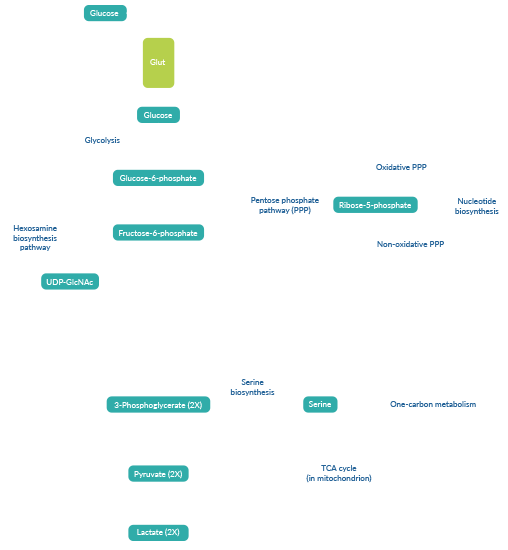
Mitochondrial Function
Mitochondrial metabolism is important in cell proliferation and tumor growth, as it supplies cells with ATP via oxidative phosphorylation, NADPH, and building blocks for macromolecule biosynthesis. Many cancer cells exhibit elevated rates of TCA cycle flux, which generates metabolites for the synthesis of nucleotides, lipids, amino acids, and heme. Anaplerotic reactions replenish TCA cycle intermediates to support continued functioning of the cycle. These include breakdown of glutamine into the TCA cycle intermediate α-ketoglutarate (α-KG) in a process known as glutaminolysis and carboxylation of pyruvate generated during glycolysis to produce oxaloacetate. Mutations in the TCA cycle enzymes succinate dehydrogenase (SDH), fumarate hydratase (FH), and isocitrate dehydrogenase 1 (IDH1) and IDH2 lead to accumulation of the oncometabolites succinate, fumarate, and 2-hydroxyglutarate (2-HG), respectively. Mitochondria also participate in one-carbon metabolism, which contributes to purine and pyrimidine nucleotide synthesis, as well as epigenetic and post-translational methylation reactions, and is commonly dysregulated in cancer.
- MitoCheck® Mitochondrial (Tissue) Isolation Kit
- MitoCheck® Complex I-V Activity Assay Kits
- Oxidative Phosphorylation Inhibitors
- α-Ketoglutarate Detection Assay Kit
- Pyruvate Assay Kit
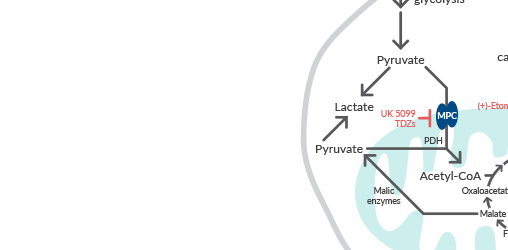
Learn more about manipulating mitochondrial fuel pathways
READ THE ARTICLE
Explore additional assay kits to study mitochondrial function
DOWNLOAD THE BROCHUREOxygen & Reactive Oxygen Species
Altered metabolic pathways in cancer cells lead to increased production of reactive oxygen species (ROS). Oxygen is reduced to superoxide (O2-) by the mitochondrial electron transport chain (ETC) or cytosolic NADPH oxidases (NOXs). Superoxide dismutase (SOD) enzymes then convert O2- to H2O2, which can act as a signaling molecule at low levels to promote cell proliferation, invasion, and tumorigenesis. At higher levels, however, ROS induce cellular damage and death. Cancer cells can buffer this increase and maintain redox homeostasis via activation of the transcription factor nuclear factor erythroid 2-related factor 2 (Nrf2), which induces expression of enzymes involved in glutathione (GSH) and NADPH production, as well as antioxidant enzymes including glutathione peroxidases (GPXs), thioredoxins (Trxs), and peroxiredoxins (Prxs).
- NOX Inhibitors
- Superoxide Dismutase Assay Kit
- Fluorescent ROS Probes
- Hydrogen Peroxide Assay Kit
- Hydrogen Peroxide Ratiometric MaxSpec® Kit
- ROS Detection Cell-Based Assay Kit (DCFDA)
- ROS Detection Cell-Based Assay Kit (DHE)
- Mitochondrial ROS Detection Assay Kit
- Nrf2 Modulators
- Nrf2 Assay Kits
- Antioxidant Assay Kit
- Glutathione Cell-Based Detection Kit (Blue Fluorescence)
- Glutathione Assay Kit
- Glutathione Peroxidase Assay Kit
- Thioredoxin Fluorometric Activity Assay Kit
Explore additional oxidative stress products in our new guide to Oxidative Stress: Markers & Detection Tools
- ROS assay kits and probes
- Antioxidant enzyme assay kits (SOD, GPXs, Trxs, and more)
Nutrient Recycling & Scavenging
In nutrient-poor conditions, cancer cells rely on recycling and scavenging pathways to break down macromolecules into building blocks that sustain survival and proliferation. Autophagy is an intracellular recycling pathway that is upregulated by nutrient stress. In these conditions, inactivation of the negative regulator mammalian target of rapamycin (mTOR) complex 1 (mTORC1) and/or activation of the positive regulator AMP-activated protein kinase (AMPK) leads to activation of the unc-51-like autophagy activating kinase 1 (ULK1) complex. This complex triggers a series of events facilitating the non-selective sequestration of cytoplasmic contents and organelles in double membrane vesicles, termed autophagosomes, that deliver their cargo to the lysosome for degradation and recycling, thereby providing building blocks to support the survival and proliferation of cancer cells.
- Autophagy-Focused Screening Library
- Autophagy/Cytotoxicity Dual Staining Kit
- mTOR Inhibitors
- AMPK Activators
- ULK1 Inhibitors
VIEW ALL:
AUTOPHAGY PRODUCTS FOR CANCER RESEARCHAUTOPHAGY PRODUCTS FOR CELL BIOLOGY RESEARCHCancer cells can obtain nutrients from the extracellular environment via scavenging mechanisms, including receptor-mediated scavenging of albumin and extracellular matrix proteins, uptake of entire live cells through entosis, and non-selective, bulk uptake via macropinocytosis. In macropinocytosis, activation of small GTPases Rac1 and Cdc42 stimulates p21-activated kinase 1 (PAK1) to induce actin polymerization and remodeling for cell surface ruffle formation. Protrusion, folding, and closure of these ruffles allows for internalization of a variety of cargo, including proteins, exosomes, and necrotic cell debris, that can be delivered to the lysosome for degradation and recycling. Like autophagy, macropinocytotsis has also been shown to be regulated by mTORC1 and AMPK, albeit in a ULK-independent manner.
All the aforementioned recycling and scavenging pathways intersect at the lysosome, where the cargoes delivered by these pathways are degraded. The breakdown products generated by this process, including amino acids, fatty acids, sugars, and nucleosides, support energy production and provide essential nutrients and intermediates for biosynthetic pathways. Disruption of lysosome function using chloroquine, hydroxychloroquine, or vacuolar ATPase (V-ATPase) inhibitors will therefore inhibit multiple nutrient recycling and scavenging pathways.
Cayman Services
CELLULAR METABOLISM SERVICES
Our cellular metabolism experts can provide a detailed view of cellular metabolic function in your experimental system via analysis of glycolysis/glycolytic flux, metabolic substrate utilization, oxidative stress, mitochondrial function and biogenesis, and more.
Learn More
Learn how Cayman's cellular metabolism experts used Seahorse XFe96 technology to examine mitochondrial fuel flexibility in a tumor-relevant model
READ THE ARTICLE